J Res Clin Med. 13:34673.
doi: 10.34172/jrcm.025.34673
Review Article
Roles of microRNAs in thalassemia: Mechanisms, biomarkers, and therapeutic implications – a narrative review
Lantip Rujito Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing, 1, * 
Tirta Wardana Data curation, Formal analysis, Investigation, Software, Visualization, Writing – review & editing, 1
Joko Mulyanto Methodology, Validation, Writing – review & editing, 1
Ita Margaretha Nainggolan Data curation, Formal analysis, Investigation, Writing – review & editing, 2
Teguh Haryo Sasongko Resources, Validation, Writing – review & editing, 3
Author information:
1Faculty of Medicine Universitas Jenderal Soedirman, Purwokerto, Indonesia
2Eijkman Research Center for Molecular Biology, National Research and Innovation Agency, Indonesia
3Faculty of Medicine, International Medical University, Kuala Lumpur, Malaysia
Abstract
Thalassemia, a kind of hemoglobin disorder, affects more than 1.5% of people worldwide. Small RNA molecules, known as microRNAs (miRNAs), are factors in controlling gene production, including hemoglobin production and disease development. This comprehensive review delves into the functions of miRNAs in the pathophysiology of thalassemia. The miRNAs miR-16 and miR-222 have been found to influence the dysregulation of erythropoiesis. These miRNAs act in impeding normal erythropoiesis, resulting in inefficient red blood cell (RBC) production and the premature demise of erythroid progenitor cells. Moreover, other miRNAs like miR-150, miR-210, and miR-485-3p regulate iron levels by pointing molecules such as transferrin receptor 1 (TfR1), ferritin, and hepcidin, which can worsen iron overload issues. Additionally, certain miRNAs have been recognized as markers for the identification of complications related to transfusions in individuals with thalassemia. The miRNAs like miR-451a, miR-20a, and miR-21 are associated with conditions such as iron overload and liver damage. Strategies involving the manipulation of the RNA molecules through mimics or inhibitors show promise in rebalancing gene activity and alleviating symptoms associated with thalassemia based on clinical trials conducted previously. The current review sheds light on how these miRNAs play a key role in modulating immune responses, and stress levels, along with organ-specific issues like heart or liver problems in patients with thalassemia. The miRNAs, including those mentioned above, impact immune cell performance, inflammation processes, and fibrosis development, contributing to the emergence of complications following transfusions. In conclusion, this comprehensive narrative review underlines the critical role of miRNAs in the pathogenesis of thalassemia. The findings illuminate their multifaceted potential as diagnostic biomarkers, therapeutic capacities, and cornerstones for developing tailored medical approaches. By harnessing these insights, clinicians and researchers may revolutionize thalassemia management, significantly improving patients’ health outcomes and quality of life.
Keywords: Erythropoiesis, Gene therapy, microRNA, Thalassemia
Copyright and License Information
© 2025 The Author(s).
This is an open access article distributed under the terms of the Creative Commons Attribution License (
http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Funding Statement
Part of this project was funded by the International Research Collaboration scheme of BLU UNSOED, Ministry of Research, Technology and Higher Education Indonesia research grant No 27.49 /UN23.37/PT.01.03/II/2023.
Introduction
Thalassemia, characterized by reduced or absent hemoglobin production, poses a significant public health issue, affecting 1.5% of the global population who carry a mutated gene in their blood.1 The condition typically requires regular blood transfusions, leading to potential complications such as excessive iron accumulation and organ damage. The mutations on the beta and alpha genes are a key point that affects the production of the globin chain, an essential element of hemoglobin.2,3 The alterations interfere normal production of hemoglobin, leading to severe anemia. Small non-coding RNA molecules, microRNAs (miRNAs), provide a crucial mechanism in post-transcriptional control of fine-tuning gene expression and might play a part in managing the genes expression involved in globin synthesis.4
Emerging research suggests that miRNAs may be possible biomarkers for predicting and tracking complications in patients with thalassemia, result from frequent blood transfusions. regulatory miRNAs have become an exciting research focus in hemoglobin disorders, with evidence hinting at their possible contributions to the pathogenesis and potential as both diagnostic markers and therapeutic targets. This paper reviewed current knowledge on the functions of miRNAs in these conditions. It was structured into sections examining miRNAs’ diverse roles, like their potential as markers of transfusion complications, possible therapeutic uses, and effects on immunity, oxidative stress, iron, cardiac and liver function, red blood cell (RBC) formation, and bone marrow. Each section summarized recent studies on how miRNAs could impact different facets of the development, worsening, and management of hemoglobin disorders.5 A great body of studies has also identified specific miRNAs, regulating gene globin synthesis in the thalassemia process. Loss of control of miR-144 can lead to the silencing of embryonic α-globin and ζ-globin gene expression in human erythroid cells by targeting the erythroid-specific transcription factor KLF1.6,7 A study designed by Lulli and colleagues revealed that microRNA-486-3p (miR-486-3p) functions as a modulator of γ-globin gene expression in human RBC precursors. Their research indicated that this regulation occurs through the alteration of BCL11A levels, a known repressor of γ-globin, suggesting its potential role in the clinical severity of thalassemia.8 Furthermore, dysregulation of miRNAs and related genes has been observed in pediatric beta-thalassemia, highlighting the utility as diagnostic and prognostic biomarkers various conditions.5 A major component of thalassemia managing involves dealing with complications from frequent blood transfusions, including iron overload and immune reactions.9,10 Recent research indicates that miRNAs could serve as biomarkers to anticipate and track transfusion-associated complications. Altering miRNAs represents a thrilling and promising tactic for advancing new thalassemia therapeutic schedules by modulating various pathways. Potential modalities include using synthetic miRNA analogs, miRNA inhibitors, and gene editing approaches.11,12 This article outlined current research on miRNAs’ involvement in thalassemia, exploring their impact on disease mechanisms and potential applications in diagnosis and treatment. The key questions we seek to address include (1) How do specific miRNAs contribute to the dysregulation of erythropoiesis and ineffective erythropoiesis observed in thalassemia? (2) What is the potential of miRNAs for predicting and monitoring transfusion-related complications in thalassemia patients? (3) How can miRNA modulation strategies be leveraged for developing novel therapeutic approaches for thalassemia? (4) What are the mechanisms by which miRNAs influence immune responses, oxidative stress, iron metabolism, and organ-specific complications, such as cardiac and liver dysfunction, in thalassemia patients? By systematically examining the available evidence, we aimed to elucidate the multifaceted roles of miRNAs in thalassemia and explore their potential as diagnostic markers, therapeutic targets, and avenues for personalized medicine to improve patient outcomes
Methods
Literature search strategy
We performed a comprehensive search on the present literature to identify relevant studies. Data was collected by conducting searches in a common database, including PubMed, Science Direct, and Google Scholar, with English language and no limitation on date publication. The search terms used were a combination of keywords related to “microRNA,” “thalassemia,” “erythropoiesis,” “iron metabolism,” “oxidative stress,” “immune response,” “cardiac function,” “liver function,” “biomarkers,” and “therapeutic targets.” Additionally, the reference lists of related review articles and included studies were manually screened to identify any additional pertinent publications.
Inclusion and exclusion criteria
Studies were included if they met requirements as follows: (1) original research articles or review articles focusing on the roles of miRNAs in thalassemia pathophysiology, diagnosis, or treatment; (2) All in vitro, in vivo, and clinical studies performed on miRNAs effects in thalassemia; (3) studies reporting the association between specific miRNAs and thalassemia aspects, such as erythropoiesis, iron metabolism, oxidative stress, immune response, cardiac or liver function, transfusion-related complications, or potential therapeutic implications. Studies were excluded if they were non-English publications, case reports, commentaries, letters to the editor, or articles not relevant to the topic of miRNAs and thalassemia.
Study selection and data extraction
Two reviewers (LR and TW) independently screened the titles and abstracts of the retrieved records to identify potentially relevant studies. Then, the full-text of articles were obtained for the selected studies and evaluated against the inclusion and exclusion criteria. Data from the included studies were extracted using a common extraction form, which captured information on the study design, participant characteristics, miRNAs investigated, key findings, and conclusions.
Data synthesis and presentation
The extracted data were synthesized qualitatively and organized into subsections based on the specific roles and mechanisms of miRNAs in thalassemia, including their effects on erythropoiesis, iron metabolism, oxidative stress, immune response, cardiac and liver function, and their potential as diagnostic indicators, as well as targets for therapeutic development. Relevant findings from the included studies were summarized in each subsection, and Table 1 was designed to provide an overview of the key miRNAs and their associated functions or implications in thalassemia.
Table 1.
List of the miRNAs along with their biological aspects including in thalassemia pathophysiology
|
miRNAs
|
Biological aspects
|
| miR-16 |
Regulating normal erythropoiesis; downregulated in beta-thalassemia leading to erythroid hyperplasia. |
| miR-451 |
Regulating normal erythropoiesis; downregulated in beta-thalassemia leading to erythroid hyperplasia. |
| miR-221 |
Upregulated in alpha-thalassemia, leading to erythroid cell apoptosis |
| miR-222 |
Upregulated in alpha-thalassemia, leading to erythroid cell apoptosis |
| miR-210 |
Suppressing erythropoiesis in alpha-thalassemia; modulating oxidative stress. |
| miR-150 |
Inhibiting proliferation and differentiation of erythroid progenitor cells; modulating immune cell function (T and B lymphocytes); potential biomarker for cardiac dysfunction |
| miR-485-3p |
Regulating iron metabolism by targeting transferrin receptor 1 (TfR1) |
| miR-20a |
Regulating iron metabolism by targeting ferritin; a potential biomarker for iron overload and liver damage |
| miR-451a |
Potential biomarker for iron overload, liver fibrosis, and hemolysis/transfusion reactions |
| miR-21 |
Potential biomarker for iron overload and liver damage |
| miR-122 |
Regulating hepcidin and iron metabolism; involved in liver function and diseases. |
| miR-27b |
Inhibiting ubiquitin-specific peptidase 4 (USP4), Shielding liver cells from the effects of the TGF-β signing route; potential therapeutic target for liver fibrosis. |
| miR-23b |
Part of the miR-23b/27b/24-1 cluster, suppresses the fibrotic of the liver. |
| miR-27b |
Part of the miR-23b/27b/24-1 cluster, suppresses the fibrotic of the liver. |
| miR-24-1 |
Part of the miR-23b/27b/24-1 cluster, suppresses the fibrotic of the liver. |
| miR-765 |
Regulating cardiomyocyte contractile function and calcium cycling |
| miR-155 |
Modulating left ventricular mass index; potential therapeutic target for heart failure; modulating immune reactions associated with transfusions |
| miR-212 |
Potential therapeutic target for pathological cardiac remodeling and heart failure |
| miR-132 |
Potential therapeutic target for pathological cardiac remodeling and heart failure |
| miR-652 |
Potential therapeutic target for pathological cardiac remodeling and heart failure |
| miR-34a |
Potential therapeutic target for dilated cardiomyopathy (sex-specific effects) |
| miR-25 |
Promoting cardiomyocyte proliferation by targeting FBXW7; a potential target for cardiac regeneration. |
| miR-30d |
Regulating cardiac function through intracellular and paracrine signaling |
| miR-126 |
Regulating angiogenesis and vascular integrity; potential therapeutic target for myocardial infarction |
| miR-146a |
Regulating angiogenesis and vascular integrity; potential therapeutic target for myocardial infarction |
| miR- let-7b |
Regulating ferroportin and iron metabolism |
| miR-155 |
Modulating immune reactions associated with transfusions |
| miR-146a |
Potential biomarker for inflammation-related complications like sepsis |
| miR-223 |
Potential biomarker for inflammation-related complications like sepsis |
| miR-92a-3p |
Associated with hemolysis/transfusion reactions |
| miR-16-5p |
Associated with hemolysis/transfusion reactions |
| miR-let-7b |
Controlling inflammation by regulating the NF-κB pathway |
| miR-155 |
Controlling inflammation by regulating the NF-κB pathway |
| miR-146a |
Controlling inflammation by regulating the NF-κB pathway |
| miR-144 |
Increased sensitivity to oxidative stress |
| miR-214 |
Upregulated in thalassemia patients with oxidative stress |
| miR-92a-3p |
Regulating gamma-globin, GSH, SOD, ROS, and cell apoptosis |
| miR-9 |
Suppressing FoxO3 and affecting ROS production |
Results
The literature search yielded a total of 375 records from the electronic databases and additional records identified through manual reference screening. After removing duplicates and screening titles and abstracts, 107 full-text articles were assessed for eligibility. Of these, 97 research papers satisfied the predefined selection parameters and were incorporated into the qualitative analysis. The comprehensive analysis noted that a wide array of miRNAs play crucial roles in various aspects of thalassemia pathophysiology as depicted in Table 1.
Discussion
The following subsections will delve into the certain pathways by which miRNAs contribute to the dysregulation of erythropoiesis, modulate iron metabolism, serve as potential biomarkers for transfusion-related complications, and offer therapeutic prospects through miRNA modulation strategies. Additionally, the intricate interplay between miRNAs and immune responses, oxidative stress, and organ-specific complications in thalassemia patients will be explored. Here, we aim to underscore the significance of miRNAs as key factors in thalassemia pathogenesis, potential diagnostic biomarkers, and promising therapeutic targets, ultimately paving the way for novel personalized medicine approaches and improved patient outcomes.
Biogenesis of miRNAs
MicroRNAs are short non-coding RNA nucleotides (18-24 nucleotides long) found in untranslated regions of mRNA, regulate gene expression at a particular base sequence in mRNA, which results in either mRNA degradation or translation inhibition, thereby influencing essential cellular activities such as cell propagation, apoptosis, and differentiation.13-15 The gene will lose activity due to binding to the target mRNA. Several studies have shown that miRNAs have multiple targets involved in several diseases associated with multiple cellular processes and play a role in tumorigenesis, cardiovascular disease, diabetes, thalassemia, and others.5,16-18 In humans, miRNAs are found in all chromosomes, except the Y chromosome. RNA Polymerase II facilitates the primary transcription of these mRNA sequences into hairpin structures called Pri-miRNAs, which are transcribed from the intronic region.19 Within the nucleus, the Drosha type of RNAse III and other associated proteins form the Dorsha complex, responsible for cleaving the pri-miRNAs into pre-miRNA. Subsequently, Exportin-5 and RanGTP facilitate the transport of pre-miRNA, a nascent miRNA transcript translocated from the nuclear compartment to the cytosolic milieu. In the cytoplasm, Dicer, an RNase III endonuclease, will process the duplex pre-miRNA into a single mature miRNA strand. The single-strand miRNA then binds to the target mRNA influencing its translation pathway and determining whether the mRNA will be translated or degraded.
Role of miRNAs in thalassemia
Thalassemia is an inherited disease characterized by impaired synthesis of the beta or alpha globin chains of hemoglobin leading to chronic hemolytic anemia,20 and an increase in the number of early erythroid precursors and erythroid progenitors, which are immature cells involved in the production of RBCs. Despite the abundance of immature cells, erythropoiesis remains ineffective, resulting in suboptimal production of mature RBCs. Although there is a large pool of proliferating immature erythroblasts, the process of their maturation into functional red cells is impaired.21
Ineffective RBC production (erythropoiesis) results from two key factors: the premature death of erythroid precursors via apoptosis and, as recent studies reveal, impaired cell differentiation. This dual problem not only worsens anemia but also triggers increased iron absorption due to the body’s compensatory process. However, the dysfunctional erythropoiesis cannot utilize this extra iron effectively, promoting an iron overload and possible organ failure.22 Research has revealed that miRNAs are pivotal regulators in the aberrant mechanisms underlying thalassemia, particularly in the disruption of normal RBC formation processes. miR-16 and miR-451 are two molecules regulating normal erythropoiesis. In beta-thalassemia, downregulation of miR-16 and miR-451 leads to erythroid hyperplasia,3,23 which is typically observed in the bone marrow biopsy of individuals with ineffective erythropoiesis, and serves as a mechanism that help maintain a relatively stable RBC count, albeit abnormal size, shape, or function.
In α-thalassemia, miR-16 and miR-451 have been shown to target the 3’ untranslated region (3’-UTR) of the α-globin mRNA by decreasing its stability and translation. The study also found that miR-221 was upregulated, leading to erythroid cell apoptosis as well as miR-222,24 while miR-210 has been implicated in suppressing erythropoiesis, which disrupted the RBCs’ formation process in α-thalassemia.25-27 Several miRNAs have been also identified as negative regulators of erythropoiesis. For example, miR-150 has the potential to inhibit the proliferation and differentiation of erythroid progenitor cells by targeting c-Myb, a transcription factor critical for erythropoiesis.26,28 Similarly, miR-221/222 has been shown to inhibit erythroid differentiation by targeting the stem cell factor receptor c-kit.29
The miR-210 has been also shown to suppress erythropoietin (EPO), a hormone stimulates erythropoiesis under hypoxia condition.30 This miRNA is up-regulated in response to hypoxia and targets the 3’-UTR of the EPO mRNA, leading to its degradation and decreased production of EPO. Another finding confirmed that upregulation of miR-210 can affect the globin gene expression and erythroid differentiation in β-thalassemia/HbE erythroid progenitor.31
Additionally, long non-coding RNAs and circular RNAs, emerging evidence points to the involvement of alternative non-coding RNA species, play a role in modulating and regulating erythropoiesis. The lncRNA erythroid differentiation regulator (ERDR1) also promote erythroid differentiation by interacting with the transcription factor GATA1, which is critical for erythropoiesis.32 miRNA as systemic biomarkers in thalassemia will be discussed further in the following subsection, while Figure 1 summarizes the miRNAs involved in various metabolic pathways or systemic effects in thalassemia patients.
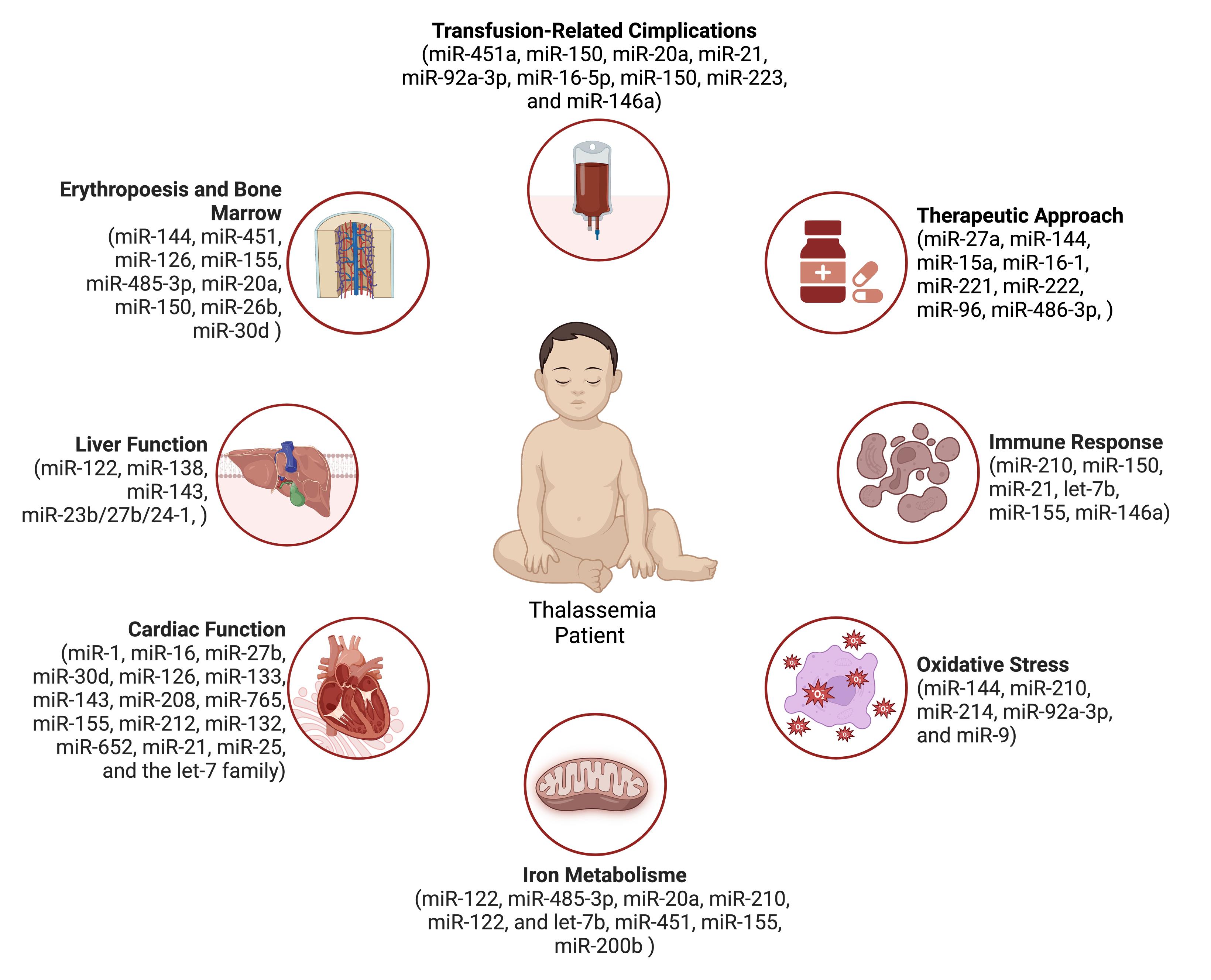
Figure 1.
miRNA and metabolism influence in Thalassemia patients
.
miRNA and metabolism influence in Thalassemia patients
miRNAs: as biomarkers for transfusion-related complications
Transfusion therapy is an essential treatment for many patients with thalassemia, but it carries the risk of various complications, including iron overload, alloimmunization, and infections.33-35 There is a pressing need to establish biomarkers with potential to predict these complications at the early stage. The development of such diagnostic indicators would allow for more pre-emptive screening for transfusion-associated complications, facilitating timely interventions and improved patient outcomes. More research is warranted to identify novel biomarkers that are accessible, rapid, and reliable for the early diagnosis of complications following blood transfusions.16
Multiple studies have also shown altered miRNA expression in thalassemia patients with complications, indicating their potential use as diagnostic biomarkers. Specific miRNAs like miR-451a, miR-20a, and miR-21 have been found upregulated in patients with iron overload and liver fibrosis.36 miR-150 may serve as a biomarker for cardiac dysfunction.37 The miRNAs have been found to influence gene expression and contribute to various pathological conditions, notably the complications arising from frequent blood transfusions.38-40 Studies have revealed associations between miRNAs like miR-451a, miR-92a-3p, miR-16-5p, and hemolysis/transfusion reactions.41-43
Additionally, miR-150, miR-223, and miR-146a may have utility as markers for inflammation-related complications like sepsis. Overall, miRNAs can be considered to accessible and non-invasive biomarkers for the early detection of transfusion-related complications in thalassemia. More research is also needed to translate these findings into clinical settings.
44-47
miRNAs have multiple benefits compared to conventional biomarkers for detecting transfusion complications, such as stability in plasma and serum, as well as enabling precise and sensitive quantification. They can also obtain through minimally invasive techniques.48 Therefore, miRNAs could enhance diagnosis, treatment, and prognosis for transfusion-related complications. However, more studies are necessary to confirm the clinical importance of miRNA for this purpose. Standardizing methods for miRNA isolation, measurement, and normalization is also essential to ensure result consistency and reproducibility. Despite many challenges, miRNAs have the remarkable potential to transform diagnosis and management of transfusion complications, ultimately improving patient outcomes.49 Following analytical and clinical validation, miRNAs may regarded as standard biomarkers for early and non-invasive detection of complications
miRNAs and immune response
Numerous studies have detected changes in miRNA expression patterns in patients with thalassemia who develop immune-mediated issues like infections, autoimmunity, and transfusion reactions. The dysregulated miRNA profiles in these patients highlight the substantial role of miRNAs in modulating immune cell function and inflammatory processes that drive the immune-related comorbidities of thalassemia. As well, the miRNAs have broad effects on multiple aspects of immune cell biology, including development, differentiation, activation, cytokine production, cell migration, and antigen presentation. By regulating these key immunological processes, miRNAs are intricately involved in orchestrating both innate and adaptive immune responses.50-53 Upregulation of miRNAs like miR-144, miR-451, and miR-503 disrupts normal erythropoiesis, leading to ineffective erythropoiesis, that triggers inflammatory responses and oxidative stress, which can impair immune cell function and promote tissue damage.4,54 Specific miRNAs have been shown to alter immune cell activity in pathological hematology patients, contributing to inflammatory and oxidative stress responses. In previous research reported that miR-210 and miR-150 can modulate the function of T and B lymphocytes, which are critical mediators of immune reactions.55 Elevated levels of miR-210 suppress the cytokine production triggered by LPS exposure like IL-6, TNF-a, Il-1β, and CCL2 by targeting NF-kB signaling.56 microRNA-150 plays a regulatory role in the maturation of natural killer cells and invariant natural killer T cells, two distinct immune cell populations with cytotoxic functions., and its dysregulation can lead to abnormal maturation and function of these innate immune cells.57 By influencing the behavior of these vital immune cell types, dysregulated expression of miRNAs like miR-210 and miR-150 may drive immunological dysfunction and inflammation-related tissue damage in thalassemia patients.58-60 Moreover, studies have demonstrated that miR-150 can govern T cell activation and development, whereas miR-21 modulates the generation of potent cytokines including IL-6 and TNF-α.61 The miR-21 in macrophages promotes an anti-inflammatory M2 phenotype polarization by targeting PDCD4, a negative regulator of IL-10.62 In addition to effects on immune cells, research indicates that miRNAs can influence the output of cytokines and chemokines, key inflammatory mediators implicated in thalassemia pathogenesis. Several studies have also demonstrated that miR-let-7b, miR-155, and miR-146a control inflammation in thalassemia by regulating the NF-κB pathway, a central coordinator of immune responses. In thalassemia, downregulation of let-7b miRNA family members (let-7b-5p, let-7i-5p) is predicted to impact signaling pathways like TGF-β, PI3K/AKT, FoxO, Hoppo, and MAPK, which are crucial for immune cells development and function.63 By modulating this critical signaling cascade, as well as downstream cytokine and chemokine production, altered expression of these miRNAs may drive aberrant inflammation and tissue damage in patients with thalassemia.64-66
Moreover, literature indicates that miR-150 and miR-155 are integral to modulating immune reactions associated with transfusions in thalassemia. These miRNAs influence B cell maturation, T cell activation, and immunoglobulin production in frequently transfused patients with thalassemia.50,67 Furthermore, miR-150 overexpression in thalassemia can disrupt B cell development, resulting in a reduced population of mature B cells and potentially compromising antibody production and immune responses to transfused blood components.63,68 A study showed that miR-150 deficiency in mice resulted in increased numbers of B-1 cells and elevated levels of natural antibodies, which could contribute to transfusion reactions.69 The elevated level of miR-155 is stored to enhanced expression of interferon-gamma (IFN-γ) and interferon beta (IFN-β), and its deregulation can lead to uncontrolled immune cell proliferation and activation of inflammatory pathways.70 The miR-155 is upregulated upon T cell activation and plays a crucial role in regulating T cell proliferation, differentiation, and cytokine production,4 in patients with leukemia and thalassemia who have aberrant T-cell activation and an exaggerated inflammatory response, potentially contributing to a transfusion-related immune reaction.71 The miR-150 and miR-155 have also been implicated in regulating immunoglobulin production by B cells, targeting the transcription factor c-Myb, which is essential for B cell development, immunoglobulin gene recombination, and activation-induced cytidine deaminase (AID) as well as Blimp-1, both involved in class-switch recombination and plasma cell differentiation. It may lead to altered immunoglobulin production and contribute to transfusion-related immune reactions.
miRNAs and oxidative stress
The oxidative stress in thalassemia is amplified by the presence of free iron radicals, heme groups, and non-heme iron. These reactive species are generated due to the disrupted balance in globin chain production, leading to an excess of unbound iron contributing to the higher oxidative state. All of these stimulate the production of excessive free radicals. Moreover, levels of antioxidant proteins are reduced in thalassemic erythrocytes. Increased levels of oxidative stress trigger a cascade of detrimental effects, including accelerated RBC destruction, impaired erythropoiesis, and dysfunction of critical organs like the heart and liver.65,72
The miRNAs have also a significant impact on oxidative stress in patients with thalassemia. Previous research has demonstrated that elevated levels of miR-144 expression correlate with a markedly heightened susceptibility to the destructive effects of oxidative stress. The downregulation of miR-144 level was also linked to elevated levels of the nuclear factor erythroid 2-related factor 2 (NRF2) and glutathione, especially in α-thalassemia. Aberrant expression levels of miR-144 could potentially contribute to the impaired capacity of RBCs to counteract oxidative stress, consequently increasing their vulnerability to premature destruction through hemolysis.73
Another study reported that oxidative stress has the potential to induce miR-210 and miR-214 upregulation in thalassemia. The potential involvement of miR-210 in oxidative stress-induced myocardial injury has been also explored. The miR-210 exerts a shielding effect against hydrogen peroxide-triggered apoptosis in cardiomyocytes by modulating the BNIP3expression, a pro-apoptotic protein that interacts with the Bcl-2 adenovirus E1B 19kDa protein.74 The miR-214 was also found associated with higher levels of reactive oxygen species (ROS), while an inverse relationship was observed between miR-214 expression and erythrocyte index including MCV, MCH, and MCHC. During Hemoglobin H disease, the higher levels of miR-214 can be detected. However, no significant difference in miR-214 expression was observed in the immature RBCs HbE/thalassemia. However, a decrease in the activating transcription factor 4 (ATF4) target was reported in both thalassemic groups. The findings suggested that the elevated expression of miR-214 was associated with both increased oxidative stress and the severity of anemia in patients with Hemoglobin H disease. This correlation was attributed to the suppression of the ATF4 by miR-214.75
Another study revealed that increased miR-92a-3p levels led to elevated γ-globin, glutathione, and superoxide dismutase expression, while reducing ROS, malondialdehyde, and cell apoptosis. Significantly, BCL11A, a regulator of hemoglobin synthesis, was recognized as a direct target of miR-92a-3p, downregulated by this microRNA.76 Regarding the activation and quantification of ROS in thalassemia patients, it is known that miR-9 in erythroid precursor controls the expression level of FoxO3, Btg1, and Cited 2. However, FoxO3-3A prevents the inhibition of erythroid cell maturation and ROS levels caused by miR-9.77
miRNAs and iron metabolism
Given that miRNAs are essential in regulating iron metabolism by targeting iron uptake, storage, and utilization genes, available evidence highlighted the miRNAs key role toward iron metabolism in patients with thalassemia. For instance, miR-122 has been shown to regulate iron metabolism by targeting hepcidin, a key regulator of iron homeostasis.78 Specific miRNAs have been demonstrated to regulate key proteins involved in iron homeostasis. In addition, miR-485-3p has the potential to regulate the expression of transferrin receptor 1 (TfR1), which facilitates cellular iron uptake. On the other hand, elevated levels of miR-485-3p suppress TfR1 expression, leading to reduced iron acquisition. In contrast, upregulated miR-20a can diminish the ferritin levels, a protein that is essential for intracellular iron storing, and further promoting iron release.2 Through modulating the expression of these critical iron handling proteins, miRNAs like miR-485-3p and miR-20a act as integral regulators of systemic and cellular iron balance. Dysregulation of these iron-related miRNAs may contribute to the iron overload observed in patients with thalassemia.79
In addition to miR-485-3p and miR-20a, other miRNAs like miR-210, miR-122, and let-7b have emerged as regulators of iron homeostasis. Also, miR-210 is increased under low oxygen and modulates several iron-related genes including TfR1, ferroportin, and hepcidin.1 Of note, Hepcidin, a hormone that inhibits iron absorption and recycling, is also regulated by miR-122. In turn, miR-let-7b regulate ferroportin, a key iron export protein. Therefore, the miRNAs comprise an iron-regulatory network through targeting critical proteins involved in systemic and cellular iron handling.79
Various factors can disrupt miRNAs regulating iron metabolism, including genetic mutations, environmental exposures, and disease states. Mutations in miRNA genes or their targets can alter miRNA expression and function. Toxins and pollutants also influence miRNA activity. Ultimately, the disease can dysregulate iron-related miRNAs like miR-20a, miR-451, miR-146a, and miR-155, contributing to anemia, inflammation, and iron overload.36,80
It is worth noting that MiR-485-3p dysregulation has a crucial role in the development of iron deficiency anemia. MiR-122 is also critical, regulating hepcidin to control iron absorption and recycling. With iron deficiency, miR-122 is also downregulated, increasing hepcidin and reducing iron uptake, respectively. In addition, MiR-122 modulates other iron genes like TfR1, ferroportin, and HO-1. By regulating multiple effectors in iron metabolism, disruption of key miRNAs like miR-485-3p and miR-122 may profoundly alter iron homeostasis in thalassemia.52 Additional miRNAs involved in iron metabolism and deficiency include miR-let-7b, miR-20a, miR-200b, and miR-185. These miRNAs modulate genes involved in iron uptake, storage, and consumption. For example, miR-let-7b and miR-20a regulate ferroportin expression, and ferritin serum levels, respectively. Through effects on key iron handling proteins, disruption of these regulatory miRNAs may alter cellular and systemic iron equilibrium.81
miRNAs and cardiac function
The major complication observed in thalassemic individuals refers to the development of congestive heart failure (HF) due to regular blood transfusions, leading to biventricular systolic dysfunction, and early mortality due to cardiomyopathy.82-84
Existing data from sequencing projects in the healthy adult heart has identified a subset of miRNAs substantially upregulated in normal cardiomyocytes. Several miRNAs, such as miR-1, coupled with miR-16 and miR-27b, have been implicated in preserving cardiac health. However, miR-30d, accompanied by miR-126, miR-133, miR-143, and miR-208, as well as the let-7 family contributing to cardiovascular diseases (CVD). Consequently, these miRNAs may hold a pivotal role in preserving normal cardiac function and in the development of CVD.85
The abundance of miR-765 leads to diminished levels of inhibitor-1, consequently impairing the contractile ability of cardiomyocytes and modulation of calcium dynamics via the protein phosphatase 1 (PP-1) signaling pathway. Down-regulation of miR-765 may serve as an additional therapeutic target to inhibit the increased PP-1 activity and improve calcium handling in failing hearts.86 Also, it has been found that the miR-155 is upregulated in patients with HF and positively correlates with left ventricular mass index, which is a prognostic marker for HF. Previous studies, using transgenic animal models, have demonstrated the clinical relevance of miR-155 involvement in HF. Importantly, knockout of miR-155 was shown to improve cardiac remodeling, potentially through targeting the tumor protein p53-inducible nuclear protein 1 (TP53INP1).87
The studies in animal models propose that anti-miR-212 may be favorable agents for the enhance the cells remodeling during HF. It is also the same with anti-miR-132 and anti-miR-562 agents. Despite the potential effects, several challenges and drawbacks associated with miRNA-based therapeutic approaches have been recognized. As an illustration, the therapeutic efficacy of anti-miR-34a has exhibited a gender-based discrepancy, demonstrating superior outcomes in females with moderate dilated cardiomyopathy compared with male counterparts.88,89
Moreover, microRNA-21 has the potential to modulate pro-fibrotic signaling toward cardiac fibroblasts and thus manage cardiac restoration. Quantitative small RNA sequencing analysis of purified myocardial cell populations also identified the highest levels of miR-21 expression within cardiomyocytes.90
Important functional interactions between myocardial infarction-associated transcript (MIAT), and miR-150, represent new regulatory mechanisms related to ischemic HF, including the Hoxa4 factor. In a recent study, it has been found that overexpressing miR-150 can alleviate the detrimental effects caused by the MIAT after a heart attack. Through genome-wide analysis of MIAT-deficient mouse hearts, they identified Hoxa4 as a novel downstream target of the MIAT/miR-150 axis. Notably, Hoxa4 was also found to be upregulated in patients with HF, underscoring the potential clinical significance of this molecular pathway.91 MicroRNA-25 was also enriched in early-stage human embryonic stem cell-originated cardiac progenitors, however, its levels steadily declined over time. Increasing the levels of miR-25 facilitated the multiplication of cardiomyocytes by downregulating the expression of F-box and WD repeat domain containing 7 (FBXW7), a gene involved in regulating cell division. Zebrafish studies have also revealed the function of miR-25 in the proliferation of cardiomyocytes, making the miR-25 as a potential molecule in cardiomyocytes rejuvenation.92 Considering the positive role of some miRNAs, such as miR-30d cardioprotective effect against HF, and they may be new therapeutic targets in this era.93
Exosome-mediated modalities have been considered strikingly optimistic strategy for cardiac function regeneration. Histological examinations revealed that in rat models with myocardial infarction (MI) subjected to treatment with alginate hydrogel vehicles encapsulating exosomes overexpressing miR-126 and miR-146a mimics exhibited diminished levels of fibrosis coupled with elevated deposition of collagen fibers. The results underscore a therapeutic potential for the treatment of MI through the regulation of angiogenesis and vascular integrity.94
miRNAs and liver function
Hepatic complications in patients with thalassemia due to iron overload include inflammation, fibrosis, and finally cirrhosis. Hepatic T2*, a non-invasive technique is utilized to evaluate the excessive accumulation of iron in the liver and heart, enabling early detection and intervention before the manifestation of overt symptoms or complications associated with iron overload.95 Liver cirrhosis is regarded as the final pathological outcome of various chronic liver diseases, with fibrosis being considered its precursor. The development of fibrotic and cirrhotic liver involves various cytokines that regulate inflammatory responses, cell proliferation, and tissue remodeling processes. While specific miRNAs modulate gene expression, influencing stellate cell activation, and matrix production. The hepatic stellate cells activation is viewed as an essential event in the development of fibrosis and is considered a crucial event during the fibrotic process.96
MicroRNA-122, a microRNA species exhibiting pronounced hepatic enrichment, has been the subject of extensive investigations delving into its biological roles and clinical relevance. It fulfills critical functions in regulating hepatic processes and has also been implicated in liver pathogenesis. Although in patients with liver fibrosis, miR-122 is downregulated, it has been shown that it is upregulated in patients with liver cirrhosis. Individuals with advanced cirrhotic liver disease exhibited a marked reduction in their serum miR-29a concentrations. Serum levels of miR-138 and miR-143 have been characterized as being indicative of the later stages of liver fibrosis, thus in early stages this marker may not be effective in the diagnosis molecules.97
Stem cells derived from adipose tissue are also regarded as a conveniently accessible source of multipotent progenitor cells with the ability to differentiate into various mesenchymal lineages. These cells have emerged as a promising therapeutic option in the field of regenerative medicine. Following 70% partial hepatectomy in rats, ASCs-miR-27b was administered. This intervention led to decreased levels of cytokines associated with inflammatory responses, upregulated hepatocyte, and other growth factors. ASCs-miR-27b administration has been shown to modulate inflammatory responses and promote liver regeneration after partial hepatectomy.98
Experimental studies demonstrated that the miR-378 has emerged as a key modulator of non-alcoholic steatohepatitis (NASH) via tumor necrosis factor (TNF) α signaling. In this regard, miR-378 is an integral part of the complex molecular pathways to induce unprompted induction of inflammatory genes, which may have important implications for the development and progression of NASH. Therefore, understanding the physiological roles of miR-378 in regulating hepatic inflammation and liver injury is important to address the complexity of the development of NASH and fibrosis.99
MicroRNA 27b inhibits ubiquitin-specific peptidase 4 (USP4), an enzyme that targets TGF-β receptor 1. It has been also recognized that liver X receptor alpha (LXRα) was found to protect liver cells from adverse stimuli and attenuate liver damage and fibrotic process. The liver X receptor confers a protective effect on hepatocytes against the detrimental impact of TGF-β through transcriptional regulation of the CB2 gene, leading to the inhibition of USP4 activity by the CB2 receptor, which stabilizes TβRI. The inhibitory role of miR-27b on USP4, a deubiquitylating enzyme for TβRI, was identified downstream of the CB2 signaling pathway.100 An intravenous delivery of miR-23b/27b/24-1 lentivirus, increasing the levels of the miR-23b/27b/24-1 cluster, was found to ameliorate hepatic fibrosis in mice. This cluster exhibited an inhibitory effect on the development of hepatic fibrosis, indicating its potential as an innovative therapeutic approach for individuals suffering from this condition.101
The miRNAs modulation: as a promising option for Thalassemia Treatment
miRNAs modulation could be achieved via strategies such as stimulators and inhibitors. The miRNA mimics are synthetic RNAs designed to imitate endogenous miRNAs. While miRNA inhibitors are antisense oligonucleotides that can bind and suppress specific miRNAs.102 By altering miRNA levels, these approaches may potentially restore balanced gene expression in thalassemic cells and improve symptoms.
Several studies have examined miRNA modulation effects on thalassemia in preclinical and clinical settings. It has been shown that miR-27a overexpression could increase fetal hemoglobin production and reduce anemia in a mouse model.103 Also, miR-144 inhibition could increase fetal hemoglobin and improve erythropoiesis. Overall, miRNA modulation is a promising therapy warranting more research. Targeting relevant miRNAs could reinstate normal gene patterns and alleviate symptoms. However, further preclinical and clinical studies are required to fully evaluate safety and efficacy.
Previous literature indicated that upregulation of miR-15a and miR-16-1 may enhance erythropoiesis in β-thalassemia patients.4,24 Similarly, increased miR-221 and miR-222 could reduce erythroid apoptosis in α-thalassemia. A study of high fetal hemoglobin individuals found that the expression of miR-15a and miR-486-3p was significantly different when compared with the control group. Other miRNAs, including miR-16-1 and miR-96, also exhibit similar expression patterns, suggesting their potential involvement in fetal hemoglobin regulation. Another study also revealed that miR-486-3p and miR-15a play significant roles in fetal hemoglobin production, together with microRNA-486-3p specifically targeting and downregulating BCL11A, a transcriptional repressor of γ-globin gene expression. The miR-15a and miR-16-1 directly inhibit MYB, a hemoglobin gene silencer. Thus, both miRNAs may have an influence on fetal hemoglobin expression and could serve as therapeutic targets.104
Limitations
While this insight delivers a comprehensive overview of the roles of miRNAs in thalassemia pathophysiology, it is important to acknowledge certain limitations. A major challenge is the efficient delivery of miRNA mimics/inhibitors to target cells and tissues. Many studies use intravenous injection, which may not effectively reach the bone marrow.103 Systemic delivery risks off-target effects and toxicity. Animal models may also not fully reflect human thalassemia complexity.105,106 Many studies use single miRNAs, which does not address the multifaceted mechanisms of thalassemia.106-108 Additionally, the lack of standardized miRNA measurement impedes result comparisons and translation.107 More extensive clinical trials are needed to evaluate the safety and efficacy of miRNA modulation strategies.
Secondly, the studies included in this review are pre-clinical investigations, conducted either in vitro or in vivo. Translating these findings to human clinical settings may pose challenges due to the inherent complexity of thalassemia disorders and the potential differences in miRNA expression and regulation between model systems and human patients. Third, most studies have focused on examining the effects of individual miRNAs or a limited number of miRNAs. However, miRNAs operate within intricate regulatory networks, and their functions may be influenced by the interplay with other miRNAs, as well as other regulatory molecules such as long non-coding RNAs and transcription factors. Investigating the combined effects of multiple miRNAs and their interactions with other regulatory elements may provide a more comprehensive understanding of their roles in thalassemia. Additionally, the methodologies used for miRNA profiling and quantification vary across studies, which can lead to inconsistencies in the reported miRNA expression levels and functional implications. Standardization of miRNA analysis techniques and data normalization approaches is crucial for ensuring the reproducibility and comparability of results.
Study Highlights
What is current knowledge?
-
MicroRNAs (miRNAs) play a critical role in the regulation of gene expression in thalassemia, particularly in erythropoiesis, iron metabolism, immune responses, and oxidative stress. They are known as promising biomarkers for diagnosing transfusion-related complications and as potential therapeutic targets to modulate disease mechanisms.
What is new here?
-
This review provides a comprehensive synthesis of recent advancements in miRNA research related to thalassemia. It highlights specific miRNAs like miR-16, miR-210, and miR-451a as pivotal players in erythropoiesis and iron metabolism. The review also discusses emerging miRNA modulation strategies, including mimics and inhibitors, that could pave the way for innovative therapeutic approaches tailored to individual patient needs.
Conclusion
This review underscores the substantial role of miRNAs in thalassemia, elucidating their regulatory mechanisms, biomarker potential, and therapeutic promise. MicroRNAs intricately modulate gene expression, impacting critical processes including erythropoiesis, iron metabolism, apoptosis, and inflammation—all central to thalassemia pathogenesis. Understanding this complex regulatory network presents exciting therapeutic targets and emphasizes the clinical utility of miRNAs as diagnostic, prognostic, and treatment response biomarkers.
Future Directions
Upon the findings presented in this review, some future directions can be considered to better understanding of miRNAs in thalassemia and translate this knowledge into clinical settings:
-
Performing large-scale clinical studies to validate the promising of miRNAs as diagnostic and prognostic molecules in thalassemia patients. These studies should aim to establish standardized protocols for miRNA quantification and normalization, ensuring the reliability and reproducibility of results across different clinical settings.
-
Exploring the feasibility of miRNA-based therapeutic approaches through pre-clinical and clinical trials. This may involve developing efficient delivery systems for miRNA mimics or inhibitors, evaluating their safety and efficacy, and investigating potential combinatorial strategies with existing therapies.
-
Integrating multi-omics data, including transcriptomics, proteomics, and metabolomics, to gain a more comprehensive understanding of the regulatory networks involving miRNAs and their functional implications in thalassemia pathogenesis.
-
Investigating the potential of miRNA-based personalized medicine approaches, where individualized miRNA profiles could be used to tailor treatment strategies and monitor therapeutic responses in thalassemia patients.
-
Exploring the potential of miRNAs as targets for gene editing technologies, such as CRISPR/Cas9, to modulate their expression and function in thalassemia disorders.
-
Designing longitudinal studies to assess the dynamic changes in miRNA expression profiles throughout thalassemia progression and in response to various therapeutic interventions.
Competing Interests
The authors state that there are no conflicts of interest regarding the publication of this manuscript. The authors have made substantial contributions to the work and agree with the manuscript’s results and conclusions. This manuscript has been read and approved by all named authors, and there are no relationships or activities that have influenced or biased the work.
Ethical Approval
Not applicable.
Acknowledgements
We extend our heartfelt gratitude to Wuri and Tina for their invaluable assistance in the development of this manuscript. Their dedication and expertise were crucial in the downloading, arranging, and filtering of numerous articles, significantly contributing to the depth and quality of this paper.
References
- Hossain MS, Raheem E, Sultana TA, Ferdous S, Nahar N, Islam S. Thalassemias in South Asia: clinical lessons learnt from Bangladesh. Orphanet J Rare Dis 2017; 12(1):93. doi: 10.1186/s13023-017-0643-z [Crossref] [ Google Scholar]
- Susanto Z, Siswandari W, Rujito L. Cd60 (GTG > GAG)/Hb Cagliari mutation was found in scanning of β-thalassemia alleles from patients of East Kalimantan, Indonesia. Mol Genet Metab Rep 2020; 22:100550. doi: 10.1016/j.ymgmr.2019.100550 [Crossref] [ Google Scholar]
- Nienhuis AW, Nathan DG. Pathophysiology and clinical manifestations of the β-thalassemias. Cold Spring Harb Perspect Med 2012; 2(12):a011726. doi: 10.1101/cshperspect.a011726 [Crossref] [ Google Scholar]
- Saki N, Abroun S, Soleimani M, Kavianpour M, Shahjahani M, Mohammadi-Asl J. MicroRNA expression in β-thalassemia and sickle cell disease: a role in the induction of fetal hemoglobin. Cell J 2016; 17(4):583-92. doi: 10.22074/cellj.2016.3808 [Crossref] [ Google Scholar]
- Wang H, Chen M, Xu S, Pan Y, Zhang Y, Huang H. Abnormal regulation of microRNAs and related genes in pediatric β-thalassemia. J Clin Lab Anal 2021; 35(9):e23945. doi: 10.1002/jcla.23945 [Crossref] [ Google Scholar]
- Fu YF, Du TT, Dong M, Zhu KY, Jing CB, Zhang Y. Mir-144 selectively regulates embryonic alpha-hemoglobin synthesis during primitive erythropoiesis. Blood 2009; 113(6):1340-9. doi: 10.1182/blood-2008-08-174854 [Crossref] [ Google Scholar]
- Penglong T, Saensuwanna A, Kocharoenwat J, Boorintaragot W, Fupongsiriphan S, Srinoun K. MiR-144 regulates hemoglobin expression in human erythroid cell line. Walailak J Sci Technol 2020; 17(11):1221-9. doi: 10.48048/wjst.2020.10712 [Crossref] [ Google Scholar]
- Lulli V, Romania P, Morsilli O, Cianciulli P, Gabbianelli M, Testa U. MicroRNA-486-3p regulates γ-globin expression in human erythroid cells by directly modulating BCL11A. PLoS One 2013; 8(4):e60436. doi: 10.1371/journal.pone.0060436 [Crossref] [ Google Scholar]
- Zhou Y, Luo J. Relationship between Iron deposition and T lymphocytes in children with β-thalassemia with haematopoietic stem cell transplantation. Front Pediatr 2022; 10:939157. doi: 10.3389/fped.2022.939157 [Crossref] [ Google Scholar]
- Keshk WA, Hablas NM, Esheba NES, Abd Elsalam SA. Crosstalk between cytokine profile, redox, and iron status in β-thalassemia: relation to frequency/duration of blood transfusion. Pediatr Hematol Oncol 2019; 36(3):151-60. doi: 10.1080/08880018.2019.1607960 [Crossref] [ Google Scholar]
- Yu AM, Jian C, Yu AH, Tu MJ. RNA therapy: are we using the right molecules?. Pharmacol Ther 2019; 196:91-104. doi: 10.1016/j.pharmthera.2018.11.011 [Crossref] [ Google Scholar]
- Kotowska-Zimmer A, Pewinska M, Olejniczak M. Artificial miRNAs as therapeutic tools: challenges and opportunities. Wiley Interdiscip Rev RNA 2021; 12(4):e1640. doi: 10.1002/wrna.1640 [Crossref] [ Google Scholar]
- Dhawan A, Scott JG, Harris AL, Buffa FM. Pan-cancer characterisation of microRNA across cancer hallmarks reveals microRNA-mediated downregulation of tumour suppressors. Nat Commun 2018; 9(1):5228. doi: 10.1038/s41467-018-07657-1 [Crossref] [ Google Scholar]
- Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP. The impact of microRNAs on protein output. Nature 2008; 455(7209):64-71. doi: 10.1038/nature07242 [Crossref] [ Google Scholar]
- Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight?. Nat Rev Genet 2008; 9(2):102-14. doi: 10.1038/nrg2290 [Crossref] [ Google Scholar]
- Wardana T, Oktriani R, Murjayanto CH, Putri DU, Anwar SL, Aryandono T. MicroRNA gene signature for predicting mechanisms in nasopharyngeal carcinoma: a case study on the potential application of circulating biomarkers. Microrna 2023; 12(1):29-44. doi: 10.2174/2211536611666220919144834 [Crossref] [ Google Scholar]
- Morán L, Cubero FJ. Extracellular vesicles in liver disease and beyond. World J Gastroenterol 2018; 24(40):4519-26. doi: 10.3748/wjg.v24.i40.4519 [Crossref] [ Google Scholar]
- Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K. Characterization of microRNAs in serum: a novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 2008; 18(10):997-1006. doi: 10.1038/cr.2008.282 [Crossref] [ Google Scholar]
- de Planell-Saguer M, Rodicio MC. Analytical aspects of microRNA in diagnostics: a review. Anal Chim Acta 2011; 699(2):134-52. doi: 10.1016/j.aca.2011.05.025 [Crossref] [ Google Scholar]
- Madan U, Bhasin H, Dewan P, Madan J. Improving ineffective erythropoiesis in thalassemia: a hope on the horizon. Cureus 2021; 13(10):e18502. doi: 10.7759/cureus.18502 [Crossref] [ Google Scholar]
- Ribeil JA, Arlet JB, Dussiot M, Moura IC, Courtois G, Hermine O. Ineffective erythropoiesis in β-thalassemia. ScientificWorldJournal 2013; 2013:394295. doi: 10.1155/2013/394295 [Crossref] [ Google Scholar]
- Cazzola M. Ineffective erythropoiesis and its treatment. Blood 2022; 139(16):2460-70. doi: 10.1182/blood.2021011045 [Crossref] [ Google Scholar]
- Kirschner MB, Kao SC, Edelman JJ, Armstrong NJ, Vallely MP, van Zandwijk N. Haemolysis during sample preparation alters microRNA content of plasma. PLoS One 2011; 6(9):e24145. doi: 10.1371/journal.pone.0024145 [Crossref] [ Google Scholar]
- Bianchi N, Zuccato C, Finotti A, Lampronti I, Borgatti M, Gambari R. Involvement of miRNA in erythroid differentiation. Epigenomics 2012; 4(1):51-65. doi: 10.2217/epi.11.104 [Crossref] [ Google Scholar]
- Sayed D, Abdellatif M. MicroRNAs in development and disease. Physiol Rev 2011; 91(3):827-87. doi: 10.1152/physrev.00006.2010 [Crossref] [ Google Scholar]
- Havelange V, Garzon R. MicroRNAs: emerging key regulators of hematopoiesis. Am J Hematol 2010; 85(12):935-42. doi: 10.1002/ajh.21863 [Crossref] [ Google Scholar]
- Listowski MA, Heger E, Bogusławska DM, Machnicka B, Kuliczkowski K, Leluk J. MicroRNAs: fine tuning of erythropoiesis. Cell Mol Biol Lett 2013; 18(1):34-46. doi: 10.2478/s11658-012-0038-z [Crossref] [ Google Scholar]
- Sun Z, Wang Y, Han X, Zhao X, Peng Y, Li Y. miR-150 inhibits terminal erythroid proliferation and differentiation. Oncotarget 2015; 6(40):43033-47. doi: 10.18632/oncotarget.5824 [Crossref] [ Google Scholar]
- Lee JY, Kim M, Heo HR, Ha KS, Han ET, Park WS. Inhibition of microRNA-221 and 222 enhances hematopoietic differentiation from human pluripotent stem cells via c-KIT upregulation. Mol Cells 2018; 41(11):971-8. doi: 10.14348/molcells.2018.0244 [Crossref] [ Google Scholar]
- Sarakul O, Vattanaviboon P, Tanaka Y, Fucharoen S, Abe Y, Svasti S. Enhanced erythroid cell differentiation in hypoxic condition is in part contributed by miR-210. Blood Cells Mol Dis 2013; 51(2):98-103. doi: 10.1016/j.bcmd.2013.03.005 [Crossref] [ Google Scholar]
- Siwaponanan P, Fucharoen S, Sirankapracha P, Winichagoon P, Umemura T, Svasti S. Elevated levels of miR-210 correlate with anemia in β-thalassemia/HbE patients. Int J Hematol 2016; 104(3):338-43. doi: 10.1007/s12185-016-2032-0 [Crossref] [ Google Scholar]
- Xu C, Shi L. Long non-coding RNAs during normal erythropoiesis. Blood Sci 2019; 1(2):137-40. doi: 10.1097/bs9.0000000000000027 [Crossref] [ Google Scholar]
- Patterson S, Singleton A, Branscomb J, Nsonwu V, Spratling R. Transfusion complications in thalassemia: patient knowledge and perspectives. Front Med (Lausanne) 2022; 9:772886. doi: 10.3389/fmed.2022.772886 [Crossref] [ Google Scholar]
- Betts M, Flight PA, Paramore LC, Tian L, Milenković D, Sheth S. Systematic literature review of the burden of disease and treatment for transfusion-dependent β-thalassemia. Clin Ther 2020;42(2):322-37.e2. doi: 10.1016/j.clinthera.2019.12.003.
- Taher AT, Radwan A, Viprakasit V. When to consider transfusion therapy for patients with non-transfusion-dependent thalassaemia. Vox Sang 2015; 108(1):1-10. doi: 10.1111/vox.12201 [Crossref] [ Google Scholar]
- Bravo Vázquez LA, Moreno Becerril MY, Mora Hernández EO, de León Carmona GG, Aguirre Padilla ME, Chakraborty S. The emerging role of microRNAs in bone diseases and their therapeutic potential. Molecules 2021; 27(1):211. doi: 10.3390/molecules27010211 [Crossref] [ Google Scholar]
- Wang H, Peng W, Ouyang X, Li W, Dai Y. Circulating microRNAs as candidate biomarkers in patients with systemic lupus erythematosus. Transl Res 2012; 160(3):198-206. doi: 10.1016/j.trsl.2012.04.002 [Crossref] [ Google Scholar]
- Peters LJ, Biessen EA, Hohl M, Weber C, van der Vorst EP, Santovito D. Small things matter: relevance of microRNAs in cardiovascular disease. Front Physiol 2020; 11:793. doi: 10.3389/fphys.2020.00793 [Crossref] [ Google Scholar]
- Gangwar RS, Rajagopalan S, Natarajan R, Deiuliis JA. Noncoding RNAs in cardiovascular disease: pathological relevance and emerging role as biomarkers and therapeutics. Am J Hypertens 2018; 31(2):150-65. doi: 10.1093/ajh/hpx197 [Crossref] [ Google Scholar]
- Ratnasari N, Lestari P, Renovaldi D, Raditya Ningsih J, Qoriansas N, Wardana T. Potential plasma biomarkers: miRNA-29c, miRNA-21, and miRNA-155 in clinical progression of hepatocellular carcinoma patients. PLoS One 2022; 17(2):e0263298. doi: 10.1371/journal.pone.0263298 [Crossref] [ Google Scholar]
- Khowawisetsut L, Vimonpatranon S, Lekmanee K, Sawasdipokin H, Srimark N, Chotivanich K. Differential effect of extracellular vesicles derived from Plasmodium falciparum-infected red blood cells on monocyte polarization. Int J Mol Sci 2023; 24(3):2631. doi: 10.3390/ijms24032631 [Crossref] [ Google Scholar]
- Mencias M, Levene M, Blighe K, Bax BE. Circulating miRNAs as biomarkers for mitochondrial neuro-gastrointestinal encephalomyopathy. Int J Mol Sci 2021; 22(7):3681. doi: 10.3390/ijms22073681 [Crossref] [ Google Scholar]
- Mackie FL, Baker BC, Beggs AD, Stodolna A, Morris RK, Kilby MD. MicroRNA changes in maternal serum from pregnancies complicated by twin-twin transfusion syndrome: a discovery study. Prenat Diagn 2019; 39(8):616-34. doi: 10.1002/pd.5475 [Crossref] [ Google Scholar]
- Shams K, Kurowska-Stolarska M, Schütte F, Burden AD, McKimmie CS, Graham GJ. MicroRNA-146 and cell trauma down-regulate expression of the psoriasis-associated atypical chemokine receptor ACKR2. J Biol Chem 2018; 293(8):3003-12. doi: 10.1074/jbc.M117.809780 [Crossref] [ Google Scholar]
- Průcha M, Černý V. Immunotherapy of sepsis. Anest Intenziv Med 2010; 21(1):23-30. [ Google Scholar]
- Osman A, Hitzler WE, Meyer CU, Landry P, Corduan A, Laffont B. Effects of pathogen reduction systems on platelet microRNAs, mRNAs, activation, and function. Platelets 2015; 26(2):154-63. doi: 10.3109/09537104.2014.898178 [Crossref] [ Google Scholar]
- Quan C, Wang M, Chen H, Zhang H. Extracellular vesicles in acute respiratory distress syndrome: recent developments from bench to bedside. Int Immunopharmacol 2021; 100:108118. doi: 10.1016/j.intimp.2021.108118 [Crossref] [ Google Scholar]
- Ajit SK. Circulating microRNAs as biomarkers, therapeutic targets, and signaling molecules. Sensors (Basel) 2012; 12(3):3359-69. doi: 10.3390/s120303359 [Crossref] [ Google Scholar]
- Precazzini F, Detassis S, Imperatori AS, Denti MA, Campomenosi P. Measurements methods for the development of microRNA-based tests for cancer diagnosis. Int J Mol Sci 2021; 22(3):1176. doi: 10.3390/ijms22031176 [Crossref] [ Google Scholar]
- Levin C, Koren A, Rebibo-Sabbah A, Levin M, Koifman N, Brenner B. Extracellular vesicle microRNA that are involved in β-thalassemia complications. Int J Mol Sci 2021; 22(18):9760. doi: 10.3390/ijms22189760 [Crossref] [ Google Scholar]
- Hojjati MT, Azarkeivan A, Pourfathollah AA, Amirizadeh N. Comparison of microRNAs mediated in reactivation of the γ-globin in β-thalassemia patients, responders and non-responders to hydroxyurea. Hemoglobin 2017; 41(2):110-5. doi: 10.1080/03630269.2017.1290651 [Crossref] [ Google Scholar]
- El-Khazragy N, Matbouly S, Hanna DH, Mahran NA, Mostafa SA, Abdelrehim BA. Circulating miRNAs and tissue iron overload in transfusion-dependent β-thalassemia major: novel predictors and follow-up guide. Ann Hematol 2021; 100(12):2909-17. doi: 10.1007/s00277-021-04639-0 [Crossref] [ Google Scholar]
- Salvi V, Gianello V, Tiberio L, Sozzani S, Bosisio D. Cytokine targeting by miRNAs in autoimmune diseases. Front Immunol 2019; 10:15. doi: 10.3389/fimmu.2019.00015 [Crossref] [ Google Scholar]
- Xu P, Palmer LE, Lechauve C, Zhao G, Yao Y, Luan J. Regulation of gene expression by miR-144/451 during mouse erythropoiesis. Blood 2019; 133(23):2518-28. doi: 10.1182/blood.2018854604 [Crossref] [ Google Scholar]
- Del Gaizo M, Sergio I, Lazzari S, Cialfi S, Pelullo M, Screpanti I. MicroRNAs as modulators of the immune response in T-cell acute lymphoblastic leukemia. Int J Mol Sci 2022; 23(2):829. doi: 10.3390/ijms23020829 [Crossref] [ Google Scholar]
- Zhang D, Cao X, Li J, Zhao G. MiR-210 inhibits NF-κB signaling pathway by targeting DR6 in osteoarthritis. Sci Rep 2015; 5:12775. doi: 10.1038/srep12775 [Crossref] [ Google Scholar]
- Mi QS, Wang J, Liu Q, Wu X, Zhou L. MicroRNA dynamic expression regulates invariant NKT cells. Cell Mol Life Sci 2021; 78(16):6003-15. doi: 10.1007/s00018-021-03895-7 [Crossref] [ Google Scholar]
- Chandan K, Gupta M, Sarwat M. Role of host and pathogen-derived microRNAs in immune regulation during infectious and inflammatory diseases. Front Immunol 2019; 10:3081. doi: 10.3389/fimmu.2019.03081 [Crossref] [ Google Scholar]
- O’Connell RM, Rao DS, Chaudhuri AA, Baltimore D. Physiological and pathological roles for microRNAs in the immune system. Nat Rev Immunol 2010; 10(2):111-22. doi: 10.1038/nri2708 [Crossref] [ Google Scholar]
- Sarma NJ, Tiriveedhi V, Ramachandran S, Crippin J, Chapman W, Mohanakumar T. Modulation of immune responses following solid organ transplantation by microRNA. Exp Mol Pathol 2012; 93(3):378-85. doi: 10.1016/j.yexmp.2012.09.020 [Crossref] [ Google Scholar]
- Doghish AS, Ismail A, El-Mahdy HA, Elkhawaga SY, Elsakka EG, Mady EA. miRNAs insights into rheumatoid arthritis: favorable and detrimental aspects of key performers. Life Sci 2023; 314:121321. doi: 10.1016/j.lfs.2022.121321 [Crossref] [ Google Scholar]
- Luo X, Ranade K, Talker R, Jallal B, Shen N, Yao Y. MicroRNA-mediated regulation of innate immune response in rheumatic diseases. Arthritis Res Ther 2013; 15(2):210. doi: 10.1186/ar4194 [Crossref] [ Google Scholar]
- Wang F, Ling L, Yu D. MicroRNAs in β-thalassemia. Am J Med Sci 2021; 362(1):5-12. doi: 10.1016/j.amjms.2021.02.011 [Crossref] [ Google Scholar]
- Pasculli E, Gadaleta RM, Arconzo M, Cariello M, Moschetta A. The role of exogenous microRNAs on human health: the plant-human trans-kingdom hypothesis. Nutrients 2024; 16(21):3658. doi: 10.3390/nu16213658 [Crossref] [ Google Scholar]
- Srinoun K, Nopparatana C, Wongchanchailert M, Fucharoen S. MiR-155 enhances phagocytic activity of β-thalassemia/HbE monocytes via targeting of BACH1. Int J Hematol 2017; 106(5):638-47. doi: 10.1007/s12185-017-2291-4 [Crossref] [ Google Scholar]
- Mattè A, Kosinski PA, Federti E, Dang L, Siciliano A, Iatcenko I. Mitapivat improves transfusion burden and reduces iron overload in thalassemic mice. Blood 2021; 138:2016. doi: 10.1182/blood-2021-153721 [Crossref] [ Google Scholar]
- Vigorito E, Perks KL, Abreu-Goodger C, Bunting S, Xiang Z, Kohlhaas S. MicroRNA-155 regulates the generation of immunoglobulin class-switched plasma cells. Immunity 2007; 27(6):847-59. doi: 10.1016/j.immuni.2007.10.009 [Crossref] [ Google Scholar]
- Das SS, Das S, Byram PK, Rahaman M, Dolai TK, Chatterjee A. MicroRNA expression patterns in HbE/β-thalassemia patients: the passwords to unlock fetal hemoglobin expression in β-hemoglobinopathies. Blood Cells Mol Dis 2021; 87:102523. doi: 10.1016/j.bcmd.2020.102523 [Crossref] [ Google Scholar]
- Bezman NA, Chakraborty T, Bender T, Lanier LL. miR-150 regulates the development of NK and iNKT cells. J Exp Med 2011; 208(13):2717-31. doi: 10.1084/jem.20111386 [Crossref] [ Google Scholar]
- Zingale VD, Gugliandolo A, Mazzon E. MiR-155: an important regulator of neuroinflammation. Int J Mol Sci 2021; 23(1):90. doi: 10.3390/ijms23010090 [Crossref] [ Google Scholar]
- Vargova K, Pesta M, Obrtlikova P, Dusilkova N, Minarik L, Vargova J. MiR-155/miR-150 network regulates progression through the disease phases of chronic lymphocytic leukemia. Blood Cancer J 2017; 7(7):e585. doi: 10.1038/bcj.2017.63 [Crossref] [ Google Scholar]
- Fibach E, Rachmilewitz EA. The role of antioxidants and iron chelators in the treatment of oxidative stress in thalassemia. Ann N Y Acad Sci 2010; 1202:10-6. doi: 10.1111/j.1749-6632.2010.05577.x [Crossref] [ Google Scholar]
- Srinoun K, Sathirapongsasuti N, Paiboonsukwong K, Sretrirutchai S, Wongchanchailert M, Fucharoen S. miR-144 regulates oxidative stress tolerance of thalassemic erythroid cell via targeting NRF2. Ann Hematol 2019; 98(9):2045-52. doi: 10.1007/s00277-019-03737-4 [Crossref] [ Google Scholar]
- Diao H, Liu B, Shi Y, Song C, Guo Z, Liu N. MicroRNA-210 alleviates oxidative stress-associated cardiomyocyte apoptosis by regulating BNIP3. Biosci Biotechnol Biochem 2017; 81(9):1712-20. doi: 10.1080/09168451.2017.1343118 [Crossref] [ Google Scholar]
- Saensuwanna A, Penglong T, Srinoun K. Upregulation of miR-214 mediates oxidative stress in Hb H disease via targeting of ATF4. Hemoglobin 2021; 45(3):197-202. doi: 10.1080/03630269.2021.1941081 [Crossref] [ Google Scholar]
- Li H, Lin R, Li H, Ou R, Wang K, Lin J. MicroRNA-92a-3p-mediated inhibition of BCL11A upregulates γ-globin expression and inhibits oxidative stress and apoptosis in erythroid precursor cells. Hematology 2022; 27(1):1152-62. doi: 10.1080/16078454.2022.2128258 [Crossref] [ Google Scholar]
- Zhang Y, Li L, Yu C, Senyuk V, Li F, Quigley JG. miR-9 upregulation leads to inhibition of erythropoiesis by repressing FoxO3. Sci Rep 2018; 8(1):6519. doi: 10.1038/s41598-018-24628-0 [Crossref] [ Google Scholar]
- Wallace DF, Crawford DH, Subramaniam VN. The control of iron homeostasis: microRNAs join the party. Gastroenterology 2011; 141(4):1520-2. doi: 10.1053/j.gastro.2011.08.018 [Crossref] [ Google Scholar]
- Muckenthaler MU, Rivella S, Hentze MW, Galy B. A red carpet for iron metabolism. Cell 2017; 168(3):344-61. doi: 10.1016/j.cell.2016.12.034 [Crossref] [ Google Scholar]
- Xu J, Linneman J, Zhong Y, Yin H, Xia Q, Kang K. MicroRNAs in pulmonary hypertension, from pathogenesis to diagnosis and treatment. Biomolecules 2022; 12(4):496. doi: 10.3390/biom12040496 [Crossref] [ Google Scholar]
- Li Y, Huang X, Wang J, Huang R, Wan D. Regulation of iron homeostasis and related diseases. Mediators Inflamm 2020; 2020:6062094. doi: 10.1155/2020/6062094 [Crossref] [ Google Scholar]
- Jabbar DA, Davison G, Muslin AJ. Getting the iron out: preventing and treating heart failure in transfusion-dependent thalassemia. Cleve Clin J Med 2007; 74(11):807-10. doi: 10.3949/ccjm.74.11.807 [Crossref] [ Google Scholar]
- Nouri N, Naderi M, Rajaie S, Dorgalaleh A, Tabibian S. Evaluation of cardiac function in patients with thalassemia intermedia. Iran J Ped Hematol Oncol 2013; 3(1):193-9. [ Google Scholar]
- Farmakis D, Triposkiadis F, Lekakis J, Parissis J. Heart failure in haemoglobinopathies: pathophysiology, clinical phenotypes, and management. Eur J Heart Fail 2017; 19(4):479-89. doi: 10.1002/ejhf.708 [Crossref] [ Google Scholar]
- Romaine SP, Tomaszewski M, Condorelli G, Samani NJ. MicroRNAs in cardiovascular disease: an introduction for clinicians. Heart 2015; 101(12):921-8. doi: 10.1136/heartjnl-2013-305402 [Crossref] [ Google Scholar]
- Cai WF, Liu GS, Lam CK, Florea S, Qian J, Zhao W. Up-regulation of micro-RNA765 in human failing hearts is associated with post-transcriptional regulation of protein phosphatase inhibitor-1 and depressed contractility. Eur J Heart Fail 2015; 17(8):782-93. doi: 10.1002/ejhf.323 [Crossref] [ Google Scholar]
- Cao RY, Li Q, Miao Y, Zhang Y, Yuan W, Fan L. The emerging role of microRNA-155 in cardiovascular diseases. Biomed Res Int 2016; 2016:9869208. doi: 10.1155/2016/9869208 [Crossref] [ Google Scholar]
- Zhou SS, Jin JP, Wang JQ, Zhang ZG, Freedman JH, Zheng Y. miRNAs in cardiovascular diseases: potential biomarkers, therapeutic targets and challenges. Acta Pharmacol Sin 2018; 39(7):1073-84. doi: 10.1038/aps.2018.30 [Crossref] [ Google Scholar]
- Bergami M, Fabin N, Cenko E, Bugiardini R, Manfrini O. MicroRNAs as potential biomarkers in coronary artery disease. Curr Top Med Chem 2023; 23(6):454-69. doi: 10.2174/1568026623666221221124530 [Crossref] [ Google Scholar]
- Ramanujam D, Schön AP, Beck C, Vaccarello P, Felician G, Dueck A. MicroRNA-21-dependent macrophage-to-fibroblast signaling determines the cardiac response to pressure overload. Circulation 2021; 143(15):1513-25. doi: 10.1161/circulationaha.120.050682 [Crossref] [ Google Scholar]
- Aonuma T, Moukette B, Kawaguchi S, Barupala NP, Sepúlveda MN, Frick K. MiR-150 attenuates maladaptive cardiac remodeling mediated by long noncoding RNA MIAT and directly represses profibrotic Hoxa4. Circ Heart Fail 2022; 15(4):e008686. doi: 10.1161/circheartfailure.121.008686 [Crossref] [ Google Scholar]
- Wang B, Xu M, Li M, Wu F, Hu S, Chen X. miR-25 promotes cardiomyocyte proliferation by targeting FBXW7. Mol Ther Nucleic Acids 2020; 19:1299-308. doi: 10.1016/j.omtn.2020.01.013 [Crossref] [ Google Scholar]
- Li J, Salvador AM, Li G, Valkov N, Ziegler O, Yeri A. Mir-30d regulates cardiac remodeling by intracellular and paracrine signaling. Circ Res 2021; 128(1):e1-23. doi: 10.1161/circresaha.120.317244 [Crossref] [ Google Scholar]
- Shafei S, Khanmohammadi M, Ghanbari H, Taghdiri Nooshabadi V, Ahmadi Tafti SH, Rabbani S. Effectiveness of exosome mediated miR-126 and miR-146a delivery on cardiac tissue regeneration. Cell Tissue Res 2022; 390(1):71-92. doi: 10.1007/s00441-022-03663-4 [Crossref] [ Google Scholar]
- Bayav M, Isiksalan Ozbulbul N, Bor O. Assessment of cardiac and liver iron overload by magnetic resonance imaging in patients with thalassemia major: short-term follow-up. J Int Med Res 2020; 48(8):300060520934260. doi: 10.1177/0300060520934260 [Crossref] [ Google Scholar]
- Zhou WC, Zhang QB, Qiao L. Pathogenesis of liver cirrhosis. World J Gastroenterol 2014; 20(23):7312-24. doi: 10.3748/wjg.v20.i23.7312 [Crossref] [ Google Scholar]
- Hu J, Xu Y, Hao J, Wang S, Li C, Meng S. MiR-122 in hepatic function and liver diseases. Protein Cell 2012; 3(5):364-71. doi: 10.1007/s13238-012-2036-3 [Crossref] [ Google Scholar]
- Chen KD, Huang KT, Lin CC, Weng WT, Hsu LW, Goto S. MicroRNA-27b enhances the hepatic regenerative properties of adipose-derived mesenchymal stem cells. Mol Ther Nucleic Acids 2016; 5(2):e285. doi: 10.1038/mtna.2015.55 [Crossref] [ Google Scholar]
- Zhang T, Hu J, Wang X, Zhao X, Li Z, Niu J. MicroRNA-378 promotes hepatic inflammation and fibrosis via modulation of the NF-κB-TNFα pathway. J Hepatol 2019; 70(1):87-96. doi: 10.1016/j.jhep.2018.08.026 [Crossref] [ Google Scholar]
- Wu HM, Kim TH, Kim A, Koo JH, Joo MS, Kim SG. Liver X receptor α-induced cannabinoid receptor 2 inhibits ubiquitin-specific peptidase 4 through miR-27b, protecting hepatocytes from TGF-β. Hepatol Commun 2019; 3(10):1373-87. doi: 10.1002/hep4.1415 [Crossref] [ Google Scholar]
- Wan LY, Peng H, Ni YR, Jiang XP, Wang JJ, Zhang YQ. The miR-23b/27b/24-1 cluster inhibits hepatic fibrosis by inactivating hepatic stellate cells. Cell Mol Gastroenterol Hepatol 2022; 13(5):1393-412. doi: 10.1016/j.jcmgh.2022.01.016 [Crossref] [ Google Scholar]
- Thomson DW, Bracken CP, Szubert JM, Goodall GJ. On measuring miRNAs after transient transfection of mimics or antisense inhibitors. PLoS One 2013; 8(1):e55214. doi: 10.1371/journal.pone.0055214 [Crossref] [ Google Scholar]
- Ma Y, Wang B, Jiang F, Wang D, Liu H, Yan Y. A feedback loop consisting of microRNA 23a/27a and the β-like globin suppressors KLF3 and SP1 regulates globin gene expression. Mol Cell Biol 2013; 33(20):3994-4007. doi: 10.1128/mcb.00623-13 [Crossref] [ Google Scholar]
- Eltaweel NH, ElKamah GY, Khairat R, Atia HA, Amr KS. Epigenetic effects toward new insights as potential therapeutic target in β-thalassemia. J Genet Eng Biotechnol 2021; 19(1):51. doi: 10.1186/s43141-021-00138-x [Crossref] [ Google Scholar]
- Fellmann C, Gowen BG, Lin PC, Doudna JA, Corn JE. Cornerstones of CRISPR-Cas in drug discovery and therapy. Nat Rev Drug Discov 2017; 16(2):89-100. doi: 10.1038/nrd.2016.238 [Crossref] [ Google Scholar]
- Cavazzana M, Antoniani C, Miccio A. Gene therapy for β-hemoglobinopathies. Mol Ther 2017; 25(5):1142-54. doi: 10.1016/j.ymthe.2017.03.024 [Crossref] [ Google Scholar]
- Sun KT, Huang YN, Palanisamy K, Chang SS, Wang IK, Wu KH. Reciprocal regulation of γ-globin expression by exo-miRNAs: relevance to γ-globin silencing in β-thalassemia major. Sci Rep 2017; 7(1):202. doi: 10.1038/s41598-017-00150-7 [Crossref] [ Google Scholar]
- Gasparello J, Fabbri E, Bianchi N, Breveglieri G, Zuccato C, Borgatti M. BCL11A mRNA targeting by miR-210: a possible network regulating γ-globin gene expression. Int J Mol Sci 2017; 18(12):2530. doi: 10.3390/ijms18122530 [Crossref] [ Google Scholar]